The
Development of a Bivocal Signature Whistle in a Bottlenose Dolphin Calf
by Elke Bojanowski

Introduction

Calf releasing air bubbles during sound production
Bottlenose dolphins (Tursiops truncatus) are well known for their outstanding acoustic capabilities, especially concerning sound production. Signals of different sound categories are used in various social contexts (for a review, see Hermann 1980; Herzing 1996), and can be combined and imitated spontaneously (e.g. Reiss & McCowan 1993). Simultaneous production of whistles and clicks occurs frequently (e.g. Lilly & Miller 1961, Evans & Prescott 1962, Caldwell & Caldwell 1966), whereas reports on simultaneous whistle production remain sparse (Kaznadei et al 1976; Markov & Ostrovskaya 1990). Processes influencing the development of the acoustic repertoire, including each individual’s specific whistle (‘signature whistle’), are largely unknown (for a review, see Sayigh 1992). In the course of a long term study on the ontogeny of acoustic signals in bottlenose dolphins, four calves were recorded throughout at least their first year of life. Focusing on signature whistles, one calf named Lemon will be dealt with in detail here, as results raised new questions on the mechanisms involved in the development of the signature whistle in bottlenose dolphins.

Methods
The ‘focal calf’ Lemon, a male born in July 1997, lived in a colony of ten dolphins housed in the Dolphin Reef, Eilat, Israel. A total of 30 hours of underwater recordings were taken between July 1997 and June 1998 with a mobile underwater camera including a hydrophone. All analysed whistles (n = 933) were assigned to the specific calf by air bubbles released simultaneously to sound production (for this method, see McCowan & Reiss 1995).

Individual whistle contours of three dolphins with high degree of similarity. From left to right: focal calf Lemon, juvenile female, and male calf.
Results & Discussion
The whistle contour first developed by the calf Lemon showed a marked similarity to the signature whistles of two other individuals of the colony. At the age of 50 days, the first bivocal whistle was recorded from Lemon. From the age of 3 months on, the majority of his whistles were bivocal (~ 70%), making them clearly distinctive from all other signature whistles within the group. Two observations make it highly improbable that morphological irregularities are responsible for this signal structure: (1) From the age of three to twelve months, about 30% of all whistles recorded from Lemon were not bivocal. (2) In two cases, another calf was recorded producing an imitation of the specific whistle, including the bivocal structure.
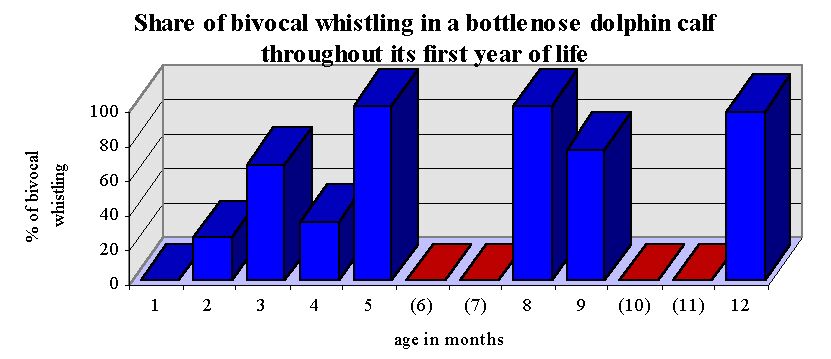
Share of bivocal whistling in the calf Lemon, given monthly throughout the first year of life. Red bars indicate that no whistles were recorded in the respective months.
The onset of bivocal whistling in the second month of life marked a decisive change in whistle structure resulting in the individual character of this animal’s signature whistle. In an additional 50 hours of underwater recordings stretching over an additional 3 year-period, no other bivocal whistles were recorded from this colony. It is therefore argued that other processes than the imitation of signals from the acoustic environment of the dolphins were likely to have influenced this development.
Summary
Within a colony of ten bottlenose dolphins, one calf developed a bivocal signature whistle. The first bivocal contour was recorded from this animal at the age of 50 days. No other bivocal whistle had been found previously in recordings taken from this group of dolphins. We conclude that the development of signature whistles in bottlenose dolphins cannot be explained by imitation of signals from the acoustic environment of the dolphins alone, but that other processes contribute.
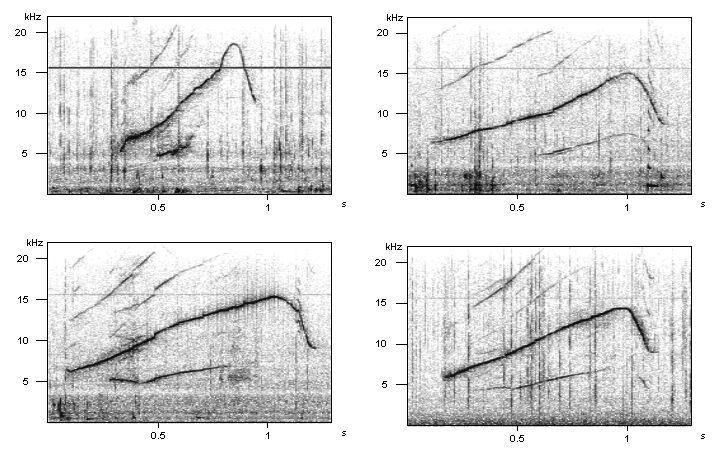
Bivocal whistles of Lemon. Top left: first bivocal whistle recorded, age: 50 days; top right and bottom left: age 6 months; bottom right: age 1 year.
References
Caldwell, M.C. & D.K. Caldwell (1966) Intraspecific Transfer of Information Via the Pulsed Sound in Captive Odontocete Cetaceans. In: Animal Sonar Systems - Biology and Bionics, R.G. Busnel (ed.), p. 879 - 936.
-
Evans, W.E. & J.H. Prescott (1962) Observations of the Sound Production Capabilities of the Bottlenosed Porpoise. Zoologica 47, p- 121 - 128.
-
Herman, L. M. & Tavolga, W. N. 1980. The Communication System of Cetaceans. In: Cetacean Behavior: Mechanisms and Functions (Ed. by L. M. Herman), pp. 149-209. New York: J. Wiley.
-
Herzing, D.L. (1996) Vocalizations and associated underwater behavior of free-ranging Atlantic spotted dolphins, Stenella frontalis, and bottlenose dolphins, Tursiops truncatus. Aquatic Mammals 22.2, p. 61 - 79.
-
Kaznadei, V.V., S.A. Kreichi & E.N. Khakhalkina (1976) Types of dolphin communication signals and their organization. Soviet Physics Acoustics 22, p. 484 - 488.
-
Lilly, J.C. & A.M Miller (1961) Sounds emitted by the bottlenose dolphin. Science 133, p. 1689 - 1693.
-
Markov, V.I. & V.M. Ostrovskaya (1990) Organization of Communication System in Tursiops truncatus Montagu. In: Sensory Abilities of Cetaceans, J. Thomas & R. Kastelein (eds.), Plenum Press, New York, p. 599 - 622.
-
McCowan, B. & D. Reiss (1995) Whistle Contour Development in Captive-Born Infant Bottlenose Dolphins (Tursiops truncatus): Role of Learning. Journal of Comparative Psychology, Vol. 109, No.3, p. 242 - 260.
-
Reiss, D. & McCowan, B. (1993) Spontaneous Vocal Mimicry and Production by Bottlenose Dolphins (Tursiops truncatus): Evidence for Vocal Learning. Journal of Comparative Psychology Vol. 107, No.3, p. 301 - 312.
-
Sayigh, L.S. (1992) Development and function of signature whistles of free-ranging bottlenose dolphins, Tursiops truncatus. Ph.D. thesis, Massachusetts Institute of Technology and Woods Hole Oceanographic Institution; 344 pages.
Source:
-
Presentation at the 14th Conference of the European Cetacean Society in Cork, Ireland, 2000.
-
PhD thesis Elke Bojanowski: Vocal Behaviour of Bottlenose Dolphins (Tursiops truncatus): Ontogeny and Contextual Use in Specific Interactions. Free University of Berlin, 2002.